4 йқ¶еҗ‘з»Ҷиғһе‘ЁжңҹиӣӢзҷҪ/ G B$ V- k5 x$ o; N1 B
з»Ҷиғһз”ҹй•ҝеҲҶиЈӮеҝ…йЎ»дҫқж¬Ўз»ҸиҝҮеҮҶеӨҮйҳ¶ж®өзҡ„й—ҙжңҹпјҲinterphaseпјүе’ҢжңүдёқеҲҶиЈӮжңҹпјҲmitosisпјүгҖӮй—ҙжңҹпјҲеҢ…жӢ¬G1гҖҒSгҖҒG2жңҹпјүзҡ„еҗ„йЎ№з”ҹе‘Ҫжҙ»еҠЁдҝқиҜҒдәҶMжңҹеҲҶиЈӮж—¶жүҖйңҖзҡ„з»ҶиғһеҶ…еҗ„жҲҗеҲҶзҡ„еӨҚеҲ¶пјҢжҜҸж¬ЎжңүдёқеҲҶиЈӮзҡ„з»“жқҹеҲ°дёӢдёҖж¬ЎжңүдёқеҲҶиЈӮзҡ„з»“жқҹжһ„жҲҗдёҖдёӘе®Ңж•ҙзҡ„з»Ҷиғһе‘ЁжңҹгҖӮз»Ҷиғһе‘Ёжңҹзҡ„иҝҗиЎҢдёҺеҗҰпјҢеҸ—жҺ§дәҺзІҫеҜҶзҡ„з»Ҷиғһе‘Ёжңҹи°ғжҺ§жңәеҲ¶гҖӮиҜҘи°ғжҺ§зі»з»ҹзҡ„ж ёеҝғжҳҜдёҖз»„з»Ҷиғһе‘Ёжңҹдҫқиө–жҖ§иӣӢзҷҪжҝҖй…¶пјҲCyclin-dependent-kinaseпјҢCDKsпјүпјҢе®ғ们еҗ„иҮӘеңЁз»Ҷиғһе‘ЁжңҹеҶ…зү№е®ҡж—¶й—ҙиў«жҝҖжҙ»пјҢйҖҡиҝҮзЈ·й…ёеҢ–еҜ№еә”зҡ„еә•зү©пјҢй©ұдҪҝз»Ҷиғһе‘Ёжңҹзҡ„е®ҢжҲҗгҖӮCDKsзҡ„ж—¶зӣёжҖ§жҝҖжҙ»дҫқиө–дәҺж—¶зӣёиЎЁиҫҫзҡ„е‘Ёжңҹзҙ пјҲcyclinпјүд»ҘеҸҠе‘Ёжңҹзҙ дҫқиө–жҖ§жҝҖй…¶жҠ‘еҲ¶еүӮпјҲcyclin-dependent kinase inhibitorsпјҢCKIпјүжҺ§еҲ¶гҖӮеҸҰеӨ–пјҢйҷӨдәҶиҝҷз§ҚжӯЈеёёз”ҹзҗҶжқЎд»¶дёӢзҡ„е‘ЁжңҹиҝӣзЁӢи°ғжҺ§пјҢеңЁй•ҝжңҹзҡ„иҝӣеҢ–иҝҮзЁӢдёӯпјҢз»Ҷиғһе»әз«ӢдәҶдёҖеҘ—дҝқиҜҒз»Ҷиғһе‘ЁжңҹдёӯйҒ—дј дҝЎжҒҜзҡ„е®Ңж•ҙжҖ§е’ҢеҮҶзЎ®жҖ§зҡ„жЈҖжҹҘжңәеҲ¶пјҢеҚіз»Ҷиғһе‘ЁжңҹжЈҖжҹҘзӮ№(checkpoint)гҖӮеҪ“з»Ҷиғһе‘ЁжңҹиҝӣзЁӢдёӯеҮәзҺ°ејӮеёёдәӢ件пјҢеҰӮDNAжҚҹдјӨжҲ–DNAеӨҚеҲ¶еҸ—йҳ»ж—¶пјҢиҝҷзұ»и°ғиҠӮжңәеҲ¶е°ұиў«жҝҖжҙ»пјҢеҸҠж—¶дёӯж–ӯз»Ҷиғһе‘Ёжңҹзҡ„иҝҗиЎҢгҖӮеҫ…DNAдҝ®еӨҚжҲ–жҺ’йҷӨдәҶж•…йҡңеҗҺпјҢз»Ҷиғһе‘ЁжңҹжүҚиғҪжҒўеӨҚиҝҗиҪ¬гҖӮ
% N C# w; j1 d
8 Z( d; ~; k; p$ V- C. |* o еңЁз»Ҷиғһзҡ„зҷҢеҸҳиҝҮзЁӢдёӯпјҢйҖҡеёёйғҪдјҙйҡҸзқҖcyclinзҡ„иҝҮеәҰиЎЁиҫҫе’ҢCKIsзҡ„зјәеӨұпјҢCDKзҡ„жҙ»жҖ§еӨұеҺ»жҺ§еҲ¶пјҢз»Ҷиғһе‘ЁжңҹеӨ„дәҺеӨұжҺ§зҠ¶жҖҒпјӣиӮҝзҳӨз»Ҷиғһзҡ„еҸҰеӨ–дёҖдёӘзү№зӮ№жҳҜз»Ҷиғһе‘ЁжңҹжЈҖжҹҘзӮ№зјәйҷ·пјҢйҖ жҲҗеҜ№з»ҶиғһжҚҹдјӨеә”зӯ”зҡ„зјәеӨұгҖӮ然иҖҢпјҢиҝҷз§Қе‘ЁжңҹжЈҖжҹҘзӮ№е…іеҚЎзҡ„зјәеӨұпјҢдҪҝеҫ—з»ҶиғһеҜ№еӨ–з•Ңзҡ„жҚҹдјӨжӣҙеҠ ж•Ҹж„ҹеҸҲиғҪиў«еә”з”ЁдәҺиӮҝзҳӨзҡ„жІ»з–—пјҢеўһеҠ ж”ҫеҢ–з–—зҡ„ж•Ҹж„ҹжҖ§гҖӮеҹәдәҺиӮҝзҳӨз»Ҷиғһзҡ„дёҠиҝ°зү№зӮ№пјҢжҒўеӨҚиӮҝзҳӨз»Ҷиғһзҡ„е‘Ёжңҹи°ғжҺ§е’ҢеҸ–ж¶ҲжЈҖжҹҘзӮ№зӯүйғҪжҲҗдёәжҪңеңЁзҡ„жҠ—иӮҝзҳӨдҪңз”Ёйқ¶зӮ№гҖӮе…·дҪ“зӯ–з•ҘеҢ…жӢ¬еҜ№CDKзҡ„зӣҙжҺҘеӮ¬еҢ–жҠ‘еҲ¶пјҢйҳ»зўҚCDKзҡ„жҝҖжҙ»пјҢе№Іжү°е‘Ёжңҹзҙ дёҺCDKзҡ„зӣёдә’дҪңз”ЁпјҢеҪұе“Қе‘Ёжңҹзҙ ж°ҙи§ЈеӨұжҙ»е’ҢжҠ‘еҲ¶з»Ҷиғһе‘ЁжңҹжЈҖжөӢзӮ№зӯүгҖӮзӣ®еүҚпјҢе·Із»ҸжңүеӨҡдёӘз»Ҷиғһе‘Ёжңҹзҡ„и°ғиҠӮеүӮиҝӣе…ҘдәҶдёҙеәҠз ”з©¶пјҢе…¶дёӯжӨҚзү©жқҘжәҗзҡ„й»„й…®зұ»зү©иҙЁflavo-piridolиғҪжҳҺжҳҫжҠ‘еҲ¶CDK1гҖҒCDK2е’ҢCDK4пјҢйҳ»зўҚз»ҶиғһйҖҡиҝҮG1/Sе’ҢG2/MжңҹжЈҖжөӢзӮ№пјҢиғҪжҠ‘еҲ¶еӨҡз§ҚиӮҝзҳӨз»Ҷиғһзҡ„з”ҹй•ҝпјҢе·Із»ҸеӨ„дәҺдёҙеәҠв…Ўжңҹз ”з©¶гҖӮеҸҰеӨ–пјҢжҳҹеһӢиӢһиҸҢзҙ пјҲStauros-porineпјүзҡ„зұ»дјјзү©UCN-01йҷӨдәҶжҠ‘еҲ¶PKCеӨ–пјҢиҝҳеҸҜзӣҙжҺҘжҠ‘еҲ¶CDK1е’ҢCDK2зҡ„жҙ»жҖ§е’Ңз»Ҷиғһе‘ЁжңҹжЈҖжөӢзӮ№жҝҖй…¶chk1зҡ„жҙ»еҢ–пјҢзӣ®еүҚжӯЈеңЁзҫҺеӣҪе’Ңж—Ҙжң¬иҝӣиЎҢдёҙеәҠв…Ўжңҹе®һйӘҢгҖӮиҝҳжңүPaullonesзұ»дјјзү©гҖҒеҳҢе‘Өйңүзҙ зұ»дјјзү©пјҲPruinesпјүзӯүйғҪеҜ№дёҚеҗҢзҡ„CDKеҲҶеӯҗжҳҫзӨәеҮәжҠ‘еҲ¶дҪңз”ЁгҖӮ5 v- @+ z3 F. _' w: G
1 Z4 y% k0 C# y% g, N( r
5 з»„иӣӢзҷҪеҺ»д№ҷй…°й…¶жҠ‘еҲ¶еүӮ(histone deacetylase inhibitorпјҢHDACi)
, y* o2 o; b T. u* A иӮҝзҳӨзҡ„еҸ‘з”ҹе’ҢиҜёеӨҡеҹәеӣ зү№еҲ«жҳҜзҷҢеҹәеӣ зҡ„ејӮеёёиЎЁиҫҫеҜҶеҲҮзӣёе…іпјҢиҖҢжҹ“иүІдҪ“з»“жһ„жҳҜи°ғжҺ§еҹәеӣ иЎЁиҫҫзҡ„йҮҚиҰҒеӣ зҙ гҖӮйҖҡеёёжғ…еҶөдёӢпјҢеҮқзј©зҡ„жҹ“иүІдҪ“дјҡжҠ‘еҲ¶еҹәеӣ зҡ„иҪ¬еҪ•пјҢиҖҢжңүиҪ¬еҪ•жҙ»жҖ§зҡ„еҹәеӣ дёҖиҲ¬дҪҚдәҺжқҫж•Јзҡ„жҹ“иүІдҪ“еҢәеҹҹгҖӮжҹ“иүІдҪ“зҡ„еҹәжң¬еҚ•дҪҚж ёе°ҸдҪ“жҳҜз”ұз»„иӣӢзҷҪпјҲHistoneпјүе’ҢDNAз»„жҲҗзҡ„пјҢе…¶дёӯз»„иӣӢзҷҪзҡ„иҪ¬еҪ•еҗҺдҝ®йҘ°пјҢеҢ…жӢ¬д№ҷй…°еҢ–гҖҒзЈ·й…ёеҢ–е’Ңз”ІеҹәеҢ–иғҪеӨҹж”№еҸҳж ёе°ҸдҪ“зҡ„й«ҳзә§з»“жһ„пјҢиҝӣиҖҢеҪұе“ҚзқҖжҹ“иүІдҪ“зҡ„й«ҳзә§з»“жһ„е’Ңеҹәеӣ зҡ„иҪ¬еҪ•и°ғжҺ§гҖӮз»ҶиғһеҶ…дёҖеҜ№еҠҹиғҪзӣёдә’жӢ®жҠ—зҡ„иӣӢзҷҪй…¶пјҢз»„иӣӢзҷҪд№ҷй…°еҹәиҪ¬з§»й…¶пјҲHistone acetyl-transferasesпјҢHATsпјүе’Ңз»„иӣӢзҷҪеҺ»д№ҷй…°й…¶пјҲHistone deace-tylasesпјҢHDACsпјүе…ұеҗҢеҶіе®ҡзқҖз»„иӣӢзҷҪзҡ„д№ҷй…°еҢ–е’ҢеҺ»д№ҷй…°еҢ–гҖӮHATеҸҜд№ҷй…°еҢ–з»„иӣӢзҷҪжң«з«ҜзўұжҖ§ж°Ёеҹәй…ёзҡ„ж°ЁеҹәпјҢдҪҝж ёе°ҸдҪ“иҲ’еұ•пјҢжҝҖжҙ»еҹәеӣ иҪ¬еҪ•гҖӮиҖҢHDACдёҺд№ӢеҠҹиғҪзӣёеҸҚпјҢжҠ‘еҲ¶еҹәеӣ иҪ¬еҪ•гҖӮ
& Q5 m# g0 A R) d5 d
- t0 O, X; T& A3 {) s4 G+ t иҝ‘дәӣе№ҙзҡ„з ”з©¶еҸ‘зҺ°HDACдҪңдёәи°ғжҺ§еҹәеӣ иҪ¬еҪ•зҡ„е…ій”®иӣӢзҷҪй…¶пјҢе…¶еҠҹиғҪејӮеёёдёҺиӮҝзҳӨзҡ„еҸ‘з”ҹе’ҢеҸ‘еұ•жңүзӣҙжҺҘе…ізі»гҖӮеҪ“HDACиҝҮеәҰиЎЁиҫҫ并被иҪ¬еҪ•еӣ еӯҗеӢҹйӣҶж—¶пјҢдјҡжҠ‘еҲ¶жҹҗдәӣеҹәеӣ зҡ„жӯЈеёёиЎЁиҫҫгҖӮиҝҷз§Қеӣ HDACжҙ»жҖ§иҝҮй«ҳеј•иө·зҡ„ејӮеёёиҪ¬еҪ•жҠ‘еҲ¶еңЁиӮҝзҳӨдёӯйқһеёёжҷ®йҒҚпјҢеӣ жӯӨHDACжҲҗдёәжҠ—иӮҝзҳӨиҚҜзү©жңҖе…·жҪңеҠӣзҡ„йқ¶зӮ№д№ӢдёҖгҖӮжҠ‘еҲ¶HDACзҡ„жҙ»жҖ§иғҪеј•иө·з»„иӣӢзҷҪй«ҳеәҰд№ҷй…°еҢ–пјҢйҮҚж–°жҝҖжҙ»жҹҗдәӣжҠ‘зҷҢеҹәеӣ зҡ„иҪ¬еҪ•е№¶еј•иө·еӨҡйЎ№дёӢжёёж•Ҳеә”пјҢеҢ…жӢ¬дҝғиҝӣиӮҝзҳӨз»ҶиғһеҲҶеҢ–гҖҒдҪҝиӮҝзҳӨз»Ҷиғһйҳ»ж»һдәҺG1жҲ–G2жңҹд»ҘеҸҠиҜұеҜјиӮҝзҳӨз»ҶиғһеҮӢдәЎпјҢд»ҺиҖҢе®һзҺ°е…¶жҠ—иӮҝзҳӨдҪңз”ЁгҖӮеҸҰеӨ–пјҢз ”з©¶еҸ‘зҺ°HDACiиҝҳиғҪжҝҖжҙ»дё»иҰҒз»„з»Үзӣёе®№жҖ§еӨҚеҗҲзү©гҖҒз»Ҷиғһй—ҙзІҳйҷ„еҲҶеӯҗICAM-1гҖҒе№Іжү°зҙ в… /в…ЎзӯүеҲҶеӯҗзҡ„иҪ¬еҪ•пјҢдҝғиҝӣе…Қз–«з»Ҷиғһзҡ„иҜҶеҲ«е’ҢжҝҖжҙ»гҖӮHDACiиҝҳиғҪжҠ‘еҲ¶зјәж°§иҜұеҜјзҡ„VEGFиЎЁиҫҫпјҢжҠ‘еҲ¶ж–°з”ҹиЎҖз®Ўз”ҹжҲҗгҖӮ8 `5 F, N, l2 Q f0 I( w4 J5 H, L
3 g% O! \5 k/ [ x/ a жқҘиҮӘзңҹиҸҢзҡ„Trichostatin AпјҲTSAпјүжҳҜеҸ‘зҺ°зҡ„йҰ–дёӘиғҪй«ҳж•ҲжҠ‘еҲ¶HDACзҡ„зҫҹиғәзұ»еӨ©з„¶дә§зү©пјҢдҪҶеӯҳеңЁзқҖеӨ©з„¶еҗ«йҮҸдҪҺгҖҒдҪ“еҶ…д»Ји°ўдёҚзЁіе®ҡзҡ„зјәзӮ№гҖӮзӣ®еүҚе·Із»Ҹжңү10еӨҡдёӘдёҚеҗҢз»“жһ„зұ»еһӢзҡ„HDACiиҝӣе…ҘдәҶв… /в…ЎжңҹдёҙеәҠиҜ•йӘҢпјҢз”ЁдәҺзҷҪиЎҖз—…е’Ңе®һдҪ“зҳӨзҡ„жІ»з–—гҖӮиҝҷдәӣиҚҜзү©еӨ§еӨҡиғҪеңЁжңүж•ҲеүӮйҮҸжҳҫзӨәеҮәиҫғеҘҪзҡ„иҖҗеҸ—жҖ§пјҢ并жҳҫзӨәеҮәжҠ—pпјҚзі–иӣӢзҷҪд»ӢеҜјзҡ„еӨҡиҚҜиҖҗиҚҜдҪңз”ЁгҖӮ& T* r' o* F. `3 b( n
, d5 n# s( e6 ?6 йқ¶еҗ‘жіӣзҙ пјҚиӣӢзҷҪй…¶дҪ“йҖҡи·Ҝ(ubiquitin-proteasome system, UPS)
. H6 ?2 |% ^6 J иӣӢзҷҪйҷҚи§Ји°ғжҺ§жҳҜз»ҶиғһдҝЎеҸ·иҪ¬еҜјзҡ„дёҖдёӘйҮҚиҰҒж–№йқўпјҢдёҺеҹәеӣ иҪ¬еҪ•ж°ҙе№ізҡ„и°ғжҺ§зӣёжҜ”пјҢиҝҷз§ҚиҪ¬еҪ•еҗҺи°ғжҺ§иҝҳиғҪдҝқиҜҒз»ҶиғһеңЁйҒҮеҲ°еӨ–з•ҢеҲәжҝҖж—¶жӣҙеҠ еҝ«йҖҹзҡ„еҒҡеҮәеҸҚеә”гҖӮUPSжҳҜзңҹж ёз»ҶиғһеҶ…дҫқиө–ATPзҡ„йқһжә¶й…¶дҪ“иӣӢзҷҪиҙЁйҷҚи§ЈйҖ”еҫ„пјҢиҙҹиҙЈи°ғжҺ§з»ҶиғһеҶ…еӨҡз§ҚиӣӢзҷҪзҡ„ж°ҙи§ЈиҝҮзЁӢпјҢе…¶дёӯеҢ…жӢ¬и®ёеӨҡи°ғиҠӮз»Ҷиғһз”ҹй•ҝгҖҒдҝЎеҸ·иҪ¬еҜјгҖҒеҹәеӣ иҪ¬еҪ•е’ҢеҮӢдәЎзҡ„йҮҚиҰҒеҲҶеӯҗгҖӮжіӣзҙ д»ӢеҜјзҡ„иӣӢзҷҪйҷҚи§ЈжҳҜдёҖдёӘеӨҚжқӮзҡ„еӨҡзә§еҸҚеә”пјҢе…¶иҝҮзЁӢдё»иҰҒжҳҜеҲ©з”Ёжіӣзҙ жҙ»еҢ–й…¶E1гҖҒжіӣзҙ з»“еҗҲй…¶E2дёҺжіӣзҙ пјҚиӣӢзҷҪиҝһжҺҘй…¶E3пјҢе°Ҷжіӣзҙ иҝһжҺҘиҮізӣ®ж ҮиӣӢзҷҪдҪңдёәж ҮиҜҶпјҢ并йҖҒиҮі20SиӣӢзҷҪй…¶дҪ“иҝӣиЎҢйҷҚи§ЈпјҢжңҖеҗҺз”ұжіӣзҙ еҲҶи§Јй…¶е°Ҷжіӣзҙ еҲҶ解并еӣһ收еҶҚеҲ©з”ЁгҖӮ
3 V" l1 M6 a! {" Z6 Q6 E" R
- w9 O& H+ D" k з”ұдәҺUPSйҖҡи·ҜдёҺиӮҝзҳӨзҡ„еҸ‘з”ҹгҖҒз”ҹй•ҝе’ҢиҪ¬з§»йғҪеҜҶеҲҮзӣёе…іпјҢиҜҘзә§иҒ”еҸҚеә”дёӯеҗ„дёӘзҺҜиҠӮйғҪжҲҗдёәжҠ—иӮҝзҳӨиҚҜзү©дҪңз”Ёзҡ„жҪңеңЁйқ¶зӮ№гҖӮдҫӢеҰӮпјҢйҖҡиҝҮйҳ»ж–ӯжіӣзҙ еҲҶеӯҗCжң«з«Ҝзҡ„и…әиӢ·й…ёеҢ–жҲ–дёҺATPеҲҶеӯҗз«һдәүз»“еҗҲзҡ„зӯ–з•ҘжқҘйҳ»зўҚжіӣзҙ зҡ„жҝҖжҙ»пјӣж №жҚ®E1дёҺE2зӣёдә’дҪңз”Ёзҡ„з»“еҗҲеҹҹзү№ејӮең°и®ҫи®ЎиғҪеӨҹе№Іжү°е…¶зӣёдә’дҪңз”Ёзҡ„е°ҸеҲҶеӯҗеҢ–еҗҲзү©йҳ»зўҚжіӣзҙ еҲҶеӯҗеңЁE1е’ҢE2д№Ӣй—ҙзҡ„дј йҖ’зӯүгҖӮе…¶дёӯзү№еҲ«еҖјеҫ—дёҖжҸҗзҡ„жҳҜйқ¶еҗ‘E3иҝһжҺҘй…¶е’Ңе…¶дёӢжёёзҡ„иӣӢзҷҪй…¶дҪ“гҖӮ3 r5 [9 q& D( u. U6 g+ A* w" ]4 o
. s9 W; }+ d" J; @; I1 I7 f E1гҖҒE2е’ҢE3жһ„жҲҗдәҶйҮ‘еӯ—еЎ”еҪўзҡ„зә§иҒ”ж”ҫеӨ§зі»з»ҹпјҢжңҖдёӢжёёзҡ„E3йҖҡиҝҮиҜҶеҲ«дёҚеҗҢеә•зү©еҶіе®ҡзқҖж•ҙдёӘжіӣзҙ еҢ–иҝҮзЁӢзҡ„зү№ејӮжҖ§гҖӮжҠ‘еҲ¶жіӣзҙ иҝһжҺҘй…¶E3зҡ„еҠҹиғҪеҸҜд»ҘйҖҡиҝҮжҠ‘еҲ¶е…¶дёҺдёҠжёёE2жҲ–дёӢжёёиӣӢзҷҪеә•зү©зҡ„дҪңз”ЁдёӨжқЎйҖ”еҫ„жқҘе®һзҺ°гҖӮзӣ®еүҚиҝҳжІЎжңүеҸ‘зҺ°й’ҲеҜ№E3е’ҢE2зӣёдә’дҪңз”Ёзҡ„жҠ‘еҲ¶еүӮпјҢдҪҶжҳҜеҜ№еҗҺиҖ…зҡ„з ”з©¶е·Із»ҸеҸ–еҫ—дәҶдёҖдәӣиҝӣеұ•пјҢе…¶дёӯжңҖе…ёеһӢзҡ„дҫӢеӯҗе°ұжҳҜMDM2еҜ№p53иӣӢзҷҪйҷҚи§Јзҡ„и°ғжҺ§гҖӮMDM2еҲҶеӯҗе…·жңүE3иҝһжҺҘй…¶жҙ»жҖ§пјҢйҖҡиҝҮжіӣзҙ еҢ–и°ғжҺ§p53зҡ„йҷҚи§ЈгҖӮNutlinsжҳҜйҰ–дёӘеҸ‘зҺ°зҡ„йҳ»зўҚp53е’ҢMDM2зӣёдә’дҪңз”Ёзҡ„е°ҸеҲҶеӯҗжҠ‘еҲ¶еүӮпјҢе…¶з©әй—ҙжһ„иұЎе’Ңp53еҲҶеӯҗдёӯдёҺMDM2дҪңз”Ёзҡ„ж°Ёеҹәй…ёж®ӢеҹәйқһеёёзӣёдјјпјҢеҸҜд»ҘдёҺp53еҲҶеӯҗз«һдәүMDM2зҡ„з»“еҗҲдҪҚзӮ№гҖӮNutlinsеңЁдҪ“еҶ…еӨ–йғҪдҪ“зҺ°еҮәжҠ—иӮҝзҳӨжҙ»жҖ§пјҢеҜ№жӯЈеёёз»„з»ҮжІЎжңүжҳҺжҳҫжҜ’жҖ§гҖӮеҸҰеӨ–дёҖдёӘе°ҸеҲҶеӯҗжҠ‘еҲ¶еүӮRITAж—©еңЁ1990е№ҙе°ұеҸ‘зҺ°е…¶жҠ—иӮҝзҳӨжҙ»жҖ§пјҢдҪҶеңЁжңҖиҝ‘жүҚеҸ‘зҺ°е…¶дҪңз”ЁжңәзҗҶжҳҜдёҺp53зҡ„Nз«Ҝз»“еҗҲпјҢйҳ»зўҚдәҶMDM2еҜ№p53зҡ„иҜҶеҲ«пјҢ并稳е®ҡp53еҲҶеӯҗNз«ҜОұ-иһәж—Ӣз»“жһ„еҹҹгҖӮ
" n0 A4 `) I& i; ?7 `$ y+ u+ N: y Z1 v+ h: K
E3иҝһжҺҘй…¶дёӢжёёзҡ„еӨҡдёӘзҺҜиҠӮпјҢеҢ…жӢ¬иӣӢзҷҪй…¶дҪ“гҖҒеҸӮдёҺжіӣзҙ жёёзҰ»еҶҚеҫӘзҺҜзҡ„йҮ‘еұһејӮиӮҪй…¶пјҲmetal-loisopeptidaseпјүд»ҘеҸҠеҜ№еӨҡиҒҡжіӣзҙ й“ҫзҡ„иҜҶеҲ«зӯүйғҪеҸҜиғҪеҪұе“ҚUPSйҖҡи·ҜгҖӮе…¶дёӯпјҢйҰ–дёӘдёҠеёӮзҡ„д»ҘUPSдёәйқ¶зӮ№зҡ„е°ҸеҲҶеӯҗжҠ‘еҲ¶еүӮbortezomibпјҲVelcadeпјҢPS-341пјүе°ұжҳҜзӣҙжҺҘжҠ‘еҲ¶дәҺиӣӢзҷҪй…¶дҪ“жҙ»жҖ§гҖӮ: O; U- S. b' B! j! z
8 y* V2 ^, P1 T" O& r( T
иҜҘеҢ–еҗҲзү©е·Іе…ҲеҗҺдәҺ2003е№ҙ5жңҲе’Ң2004е№ҙ4жңҲиў«зҫҺеӣҪFDAе’Ң欧зӣҹиҚҜе“Ғе®ЎиҜ„з®ЎзҗҶеұҖпјҲEuropean Agency for the Evaluation of Medicinal Products пјҢEMEAпјүжү№еҮҶз”ЁдәҺеӨҚеҸ‘жҖ§е’ҢйҡҫжІ»жҖ§еӨҡеҸ‘жҖ§йӘЁй«“зҳӨзҡ„жІ»з–—гҖӮжңҖиҝ‘FDAеҸҲжү№еҮҶе…¶дҪңдёәдёҖзәҝиҚҜзү©з”ЁдәҺе·ІжҺҘеҸ—иҝҮиҮіе°‘дёҖж¬ЎжІ»з–—зҡ„еӨҡеҸ‘жҖ§йӘЁй«“зҳӨжӮЈиҖ…гҖӮиҝӣдёҖжӯҘеңЁиЎҖж¶Ізі»з»ҹиӮҝзҳӨгҖҒе®һдҪ“зҳӨд»ҘеҸҠйқһйңҚеҘҮйҮ‘ж·Ӣе·ҙзҳӨдёӯзҡ„еә”з”ЁжӯЈеңЁз ”究дёӯгҖӮ. |$ K5 c+ Q3 W1 V' v* d( w: I
& s% B$ o4 N4 {$ u% \0 `" T u. x+ [7 йқ¶еҗ‘DNAжҚҹдјӨдҝ®еӨҚзі»з»ҹ* V* x% v u) d
йҷӨдәҶдёҠиҝ°зӣҙжҺҘеҪұе“ҚиӮҝзҳӨз»Ҷиғһеўһж®–и°ғжҺ§зҡ„дҝЎеҸ·йҖҡи·ҜеӨ–пјҢзү№ејӮйқ¶еҗ‘DNAжҚҹдјӨдҝ®еӨҚйҖҡи·Ҝдёӯзҡ„дёҖдәӣе…ій”®еҲҶеӯҗд№ҹжҲҗдёәжҠ—иӮҝзҳӨиҚҜзү©з ”еҸ‘зҡ„дёҖдёӘйҮҚиҰҒж–№еҗ‘гҖӮеҫҲеӨҡдј з»ҹзҡ„жҠ—иӮҝзҳӨиҚҜзү©еҢ…жӢ¬зғ·еҢ–еүӮгҖҒDNAеөҢе…ҘеүӮгҖҒжӢ“жү‘ејӮжһ„й…¶жҠ‘еҲ¶еүӮгҖҒжҠ—д»Ји°ўзү©зӯүйғҪжҳҜйҖҡиҝҮзӣҙжҺҘжҲ–й—ҙжҺҘйҖ жҲҗдёҚеҗҢеҪўејҸзҡ„DNAжҚҹдјӨжқҘе®һзҺ°е…¶жҠ—иӮҝзҳӨдҪңз”ЁгҖӮеңЁеӨ–з•ҢжҚҹдјӨзҡ„еҲәжҝҖдёӢпјҢз»ҶиғһиғҪеҗҜеҠЁ6жқЎдҝ®еӨҚйҖҡи·ҜжқҘеҲҶеҲ«еә”еҜ№дёҚеҗҢзұ»еһӢзҡ„жҚҹдјӨпјҡв‘ зӣҙжҺҘдҝ®еӨҚпјҲdirect repairпјҢDRпјүйҖҡи·Ҝдҝ®еӨҚO6пјҚзғ·еҹәйёҹеҳҢе•үеј•иө·зҡ„жҚҹдјӨпјӣв‘ЎзўұеҹәеҲҮйҷӨдҝ®еӨҚпјҲbase excision repairпјҢBERпјүй’ҲеҜ№еӣ ж°§еҢ–иҝҳеҺҹжҲ–зғ·еҹәеҢ–еј•иө·зҡ„зўұеҹәжҚҹдјӨпјӣв‘ўж ёиӢ·й…ёеҲҮйҷӨдҝ®еӨҚпјҲnucleotide excision repairпјҢNERпјүдҝ®еӨҚеӣ иҫҗе°„гҖҒеҢ–еӯҰиҚҜзү©жҲ–иӣӢзҷҪпјҚDNAдәӨиҒ”еј•иө·зҡ„ж ёиӢ·й…ёж°ҙе№ізҡ„жҚҹдјӨпјӣв‘Јзўұеҹәй”ҷй…Қдҝ®еӨҚпјҲmismatch repairпјҢMMRпјүзә жӯЈзўұеҹәй”ҷй…Қпјӣв‘ӨеҗҢжәҗйҮҚз»„дҝ®еӨҚпјҲhomologous repairпјҢHRпјүпјӣв‘ҘйқһеҗҢжәҗзҡ„жң«з«ҜиҝһжҺҘпјҲnon-homologous end-joiningпјҢNHEJпјүйҖҡи·ҜпјҢе…¶дёӯеҗҺдёӨжқЎйҖҡи·Ҝдё“й—Ёдҝ®еӨҚDNAеҸҢй“ҫж–ӯиЈӮпјҲDNA double strand breaksпјҢDSBsпјүгҖӮиҝҷдәӣйҖҡи·Ҝзҡ„жҝҖжҙ»еҫҖеҫҖеүҠејұдәҶеҢ–з–—иҚҜзҡ„жҠ—иӮҝзҳӨж•ҲжһңпјҢжҲҗдёәдә§з”ҹиҖҗиҚҜзҡ„дёҖдёӘйҮҚиҰҒеӣ зҙ гҖӮ
" Z, m8 A- ?9 P* Z! k& G+ c5 a
" _( O G, q. t зӣ®еүҚпјҢй’ҲеҜ№дёҠиҝ°еӨҡжқЎйҖҡи·ҜдёӯдёҖдәӣе…ій”®еҲҶеӯҗзҡ„жҠ‘еҲ¶еүӮз ”з©¶йғҪжңүдёҚеҗҢзЁӢеәҰзҡ„иҝӣеұ•гҖӮеҰӮO6-иӢ„еҹәйёҹеҳҢе‘Ө( O6-BG)еҸҠе…¶еӨҡдёӘиЎҚз”ҹзү©иғҪзү№ејӮжҠ‘еҲ¶DRйҖҡи·ҜдёӯO6пјҚзғ·еҹәйёҹеҳҢе•үпјҚDNA зғ·еҹәиҪ¬з§»й…¶пјҲAGTпјүпјҢзӣ®еүҚжӯЈеңЁдҪңдёәиҖҗиҚҜйҖҶиҪ¬еүӮдёҺз”ІеҹәеҢ–еүӮеҗҲз”ЁиҝӣиЎҢдёҙеәҠиҜ•йӘҢгҖӮиҝҳжңүйқ¶еҗ‘жҠ‘еҲ¶BERйҖҡи·Ҝдёӯзҡ„еӨҡиҒҡ(ADP-ж ёзі–)иҒҡеҗҲй…¶пјҲpoly[ADP-ribose]polymeraseпјҢPARPпјүзҡ„AG014699гҖҒNU1025гҖҒAG14361е’ҢINO-1001еҲҶеҲ«еӨ„дәҺв… пҪһв…ўжңҹдёҙеәҠе®һйӘҢгҖӮиҝҷдәӣжҠ‘еҲ¶еүӮдёҺзғ·еҢ–еүӮгҖҒжӢ“жү‘ејӮжһ„й…¶IжҠ‘еҲ¶еүӮгҖҒйЎәй“ӮжҲ–ОіпјҚе°„зәҝиҒ”еҗҲз”ЁдәҺиӮҝзҳӨз»ҶиғһжҲ–иҖ…е°Ҹйј з§»жӨҚзҳӨпјҢиғҪеҲҶеҲ«еўһеҠ еҗҺиҖ…еј•иө·зҡ„DNAжҚҹдјӨгҖҒз»Ҷиғһз”ҹй•ҝжҠ‘еҲ¶е’ҢиҚ·зҳӨе°Ҹйј зҡ„еӯҳжҙ»ж—¶й—ҙгҖӮзҺ°д»ҠпјҢе…іжіЁжңҖеӨҡзҡ„жҳҜдҝ®еӨҚDSBsзҡ„HRе’ҢNHEJйҖҡи·ҜпјҢеҲҶеҲ«еңЁиҝҷдёӨжқЎйҖҡи·Ҝдёӯжү®жј”е…ій”®и§’иүІзҡ„ATMе’ҢDNA-PKйғҪжҳҜPI3KжҝҖ酶家ж—ҸжҲҗе‘ҳпјҢеүҚж–Үе·Із»ҸжҸҗеҲ°зҡ„wortmanninгҖҒLY294002е’Ңе’–е•Ўеӣ зӯүPI3K家ж—Ҹзҡ„е№ҝи°ұжҠ‘еҲ¶еүӮйғҪиғҪйҖҡиҝҮжҠ‘еҲ¶иҝҷдёӨжқЎдҝ®еӨҚйҖҡи·Ҝзҡ„жҙ»жҖ§иө·еҲ°ж”ҫеҢ–з–—еўһж•Ҹзҡ„дҪңз”ЁгҖӮеҸҰеӨ–пјҢиҝҳжңүATPзҡ„з«һдәүжҠ‘еҲ¶еүӮKU-55933иғҪжҳҫи‘—жҠ‘еҲ¶ATMжҙ»жҖ§пјҢдё”дҪңз”ЁжҳҺжҳҫејәдәҺеҜ№PI3K家ж—Ҹе…¶е®ғжҲҗе‘ҳзҡ„жҠ‘еҲ¶ж•ҲжһңпјҢиғҪеўһејәATMй«ҳиЎЁиҫҫз»Ҷиғһж ӘеҜ№зҰ»еӯҗиҫҗе°„е’ҢVP16гҖҒйҳҝйңүзҙ гҖҒе–ңж ‘зўұзӯүйҖ жҲҗDSBsиҚҜзү©зҡ„ж•Ҹж„ҹжҖ§гҖӮDNA-PKзҡ„йҖүжӢ©жҖ§жҠ‘еҲ¶еүӮIC87361гҖҒNu7026гҖҒVanillinе’ҢSU117582д№ҹжңүзӣёдјјзҡ„дҪңз”ЁгҖӮ
7 q5 M2 v" j, ^% _; G+ U
0 B( Y! }7 l6 e- Y1 f$ ~з»“иҜӯ& h# R* x' t8 n* v% t" ^, y5 N
еӨҡдёӘеҲҶеӯҗйқ¶еҗ‘иҚҜзү©зҡ„жҲҗеҠҹдёҠеёӮдёәж–°дёҖд»ЈжҠ—иӮҝзҳӨиҚҜзү©зҡ„з ”еҸ‘жҸҗдҫӣдәҶд№җи§Ӯзҡ„еүҚжҷҜгҖӮдҪҶжҳҜпјҢеҝ…йңҖзңӢеҲ°йқ¶еҗ‘дҝЎеҸ·йҖҡи·ҜжҠ‘еҲ¶еүӮеҸӘжңүеңЁиҜҘдҝЎеҸ·йҖҡи·Ҝй«ҳеәҰжҝҖжҙ»зҡ„иӮҝзҳӨдёҠжүҚдә§з”ҹжӣҙеҘҪең°з–—ж•ҲпјҢеӣ жӯӨеҗҲзҗҶзҡ„дёҙеәҠи®ҫи®ЎжҳҜдҪ“зҺ°е…¶з–—ж•Ҳзҡ„еҝ…иҰҒеүҚжҸҗгҖӮеҸӘжңүеңЁеҗҲзҗҶзҡ„з ”з©¶зӯ–з•ҘгҖҒе…Ёйқўзҡ„дёҙеәҠеүҚиҜ„д»·е’Ң- k. A3 ]% K6 A
' M( {. ~: ]3 \0 [7 V3 _- uдёҘи°ЁеҗҲзҗҶзҡ„дёҙеәҠиҜ•йӘҢзҡ„е…ұеҗҢй…ҚеҗҲдёӢжүҚиғҪејҖеҸ‘еҮәжӣҙеӨҡжңүеёҢжңӣзҡ„йқ¶зӮ№зү№ејӮжҖ§жҠ—иӮҝзҳӨж–°иҚҜгҖӮ |


 иӮәзҷҢж–°иҚҜжқҘиўӯпјҢзӘҒз ҙиҖҗиҚҜеӣ°еўғпјҢиҝ‘еҚҠжӮЈ
дҪңиҖ…пјҡseacat
ZidesamtinibпјҲNVL-520пјүжҳҜжӯЈеңЁз ”еҸ‘зҡ„ж–°дёҖд»ЈROS1йқ¶еҗ‘иҚҜпјҢеҸҜжңүж•Ҳе…ӢеҲ¶еҗ„з§Қ
иӮәзҷҢж–°иҚҜжқҘиўӯпјҢзӘҒз ҙиҖҗиҚҜеӣ°еўғпјҢиҝ‘еҚҠжӮЈ
дҪңиҖ…пјҡseacat
ZidesamtinibпјҲNVL-520пјүжҳҜжӯЈеңЁз ”еҸ‘зҡ„ж–°дёҖд»ЈROS1йқ¶еҗ‘иҚҜпјҢеҸҜжңүж•Ҳе…ӢеҲ¶еҗ„з§Қ
 еҗ„дҪҚжңӢеҸӢпјҢжҲ‘зҲёзҲёжІЎжңүйқ¶еҗ‘иҚҜпјҢpdlеҸӘ
еҗ„дҪҚжңӢеҸӢпјҢжҲ‘зҲёзҲёжІЎжңүйқ¶еҗ‘иҚҜпјҢpdlеҸӘжңүзҷҫеҲҶд№Ӣ5пјҢиҝҷдёӘеҢ»жІ»зҺҮй«ҳдёҚй«ҳпјҢжҲ‘зҲёзҲёжҳҜиӮәи…әзҷҢпјҢеҜЎ
еҗ„дҪҚжңӢеҸӢпјҢжҲ‘зҲёзҲёжІЎжңүйқ¶еҗ‘иҚҜпјҢpdlеҸӘ
еҗ„дҪҚжңӢеҸӢпјҢжҲ‘зҲёзҲёжІЎжңүйқ¶еҗ‘иҚҜпјҢpdlеҸӘжңүзҷҫеҲҶд№Ӣ5пјҢиҝҷдёӘеҢ»жІ»зҺҮй«ҳдёҚй«ҳпјҢжҲ‘зҲёзҲёжҳҜиӮәи…әзҷҢпјҢеҜЎ
 жұӮеҠ©еҚ•е…Қз–«зәіжӯҰеҲ©з”ұжІ»з–—7дёӘжңҲеҮәзҺ°ејӮ
з—…еҸӢ们еӨ§е®¶еҘҪеӨ§еҚҠе№ҙжІЎйңІйқўдәҶпјҢе·ҰиӮәе…ЁеҲҮеҗҺпјҢе…Қ疫治疗马дёҠж»ЎдёҖе№ҙдәҶпјҢеҚҒжңҲд»ҪCTжҳҫзӨәжңү17mm
жұӮеҠ©еҚ•е…Қз–«зәіжӯҰеҲ©з”ұжІ»з–—7дёӘжңҲеҮәзҺ°ејӮ
з—…еҸӢ们еӨ§е®¶еҘҪеӨ§еҚҠе№ҙжІЎйңІйқўдәҶпјҢе·ҰиӮәе…ЁеҲҮеҗҺпјҢе…Қ疫治疗马дёҠж»ЎдёҖе№ҙдәҶпјҢеҚҒжңҲд»ҪCTжҳҫзӨәжңү17mm
 еҗ„дҪҚеӨ§е’–пјҢиҜ·ж•ҷеҘҘиҘҝиҖҗиҚҜеҗҺеҰӮдҪ•жІ»з–—-
1.иӮәи…әзҷҢжҷҡжңҹпјҢеҘіпјҢ54еІҒпјҢ2024е№ҙ10жңҲзЎ®иҜҠиӮәи…әзҷҢжҷҡжңҹи„‘иҪ¬з§»еҒҡдәҶејҖйў…жүӢжңҜ(и„‘иҪ¬з§»зҳӨдёә3.4
еҗ„дҪҚеӨ§е’–пјҢиҜ·ж•ҷеҘҘиҘҝиҖҗиҚҜеҗҺеҰӮдҪ•жІ»з–—-
1.иӮәи…әзҷҢжҷҡжңҹпјҢеҘіпјҢ54еІҒпјҢ2024е№ҙ10жңҲзЎ®иҜҠиӮәи…әзҷҢжҷҡжңҹи„‘иҪ¬з§»еҒҡдәҶејҖйў…жүӢжңҜ(и„‘иҪ¬з§»зҳӨдёә3.4
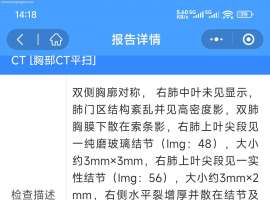 е·Із»ҸеҢ–з–—4ж¬ЎпјҢжҳҜеҗҰ继з»ӯ
жң¬дәәзҲёзҲёжҷҡжңҹиӮәзҷҢпјҲиғёиҶңиҪ¬з§»пјүпјҢеӣ еүҚжңҹеҠ ејәCTжңӘжҳҫзӨәеҮәиғёиҶңиҪ¬з§»пјҢеҢ»з”ҹз»ҷеҒҡдәҶжүӢжңҜпјҢеҒҡ
е·Із»ҸеҢ–з–—4ж¬ЎпјҢжҳҜеҗҰ继з»ӯ
жң¬дәәзҲёзҲёжҷҡжңҹиӮәзҷҢпјҲиғёиҶңиҪ¬з§»пјүпјҢеӣ еүҚжңҹеҠ ејәCTжңӘжҳҫзӨәеҮәиғёиҶңиҪ¬з§»пјҢеҢ»з”ҹз»ҷеҒҡдәҶжүӢжңҜпјҢеҒҡ





 4 q% M( k9 Y# O A* o" e
4 q% M( k9 Y# O A* o" e жҳҫиә«еҚЎ
жҳҫиә«еҚЎ